Contributed by Rosario Gulino, Vincenzo Perciavalle and Massimo Gulisan
Dept of Physiological Sciences, Univ of Catania, Viale Andrea Doria, 6-I95125 Catania ITALY
A promising approach for central nervous system (CNS) repair consists in the activation of endogenous neural precursor cells (NPCs), but this process is less efficient in the spinal cord (SC) following a spinal cord injury (SCI). Another process promoting a functional restoration after SCI consists in the reorganization of spared pathways by mechanisms involving the modulation of synaptic efficacy.[1,2] Sonic hedgehog (Shh), Notch-1 and Numb are involved in the stem cell functioning [3] and, additionally, Notch-1 has a role as modulator of synaptic plasticity. [4,5] However, little is known about the role of these proteins in the adult SC after removal of motoneurons.
In this study, we injected Cholera toxin- B saporin (CTB-SAP, Cat. #IT-14) into the gastrocnemius muscle to induce a mild depletion (about 30%; Fig. 1) of motoneurons within the lumbar SC of adult mice and analyzed the expression of Choline acetyltransferase (ChAT), Synapsin-I, Shh, Notch-1 and Numb proteins, by western blotting. The functional outcome of the lesion, as well as the possible recovery of locomotion, were monitored by grid walk and rotarod tests.
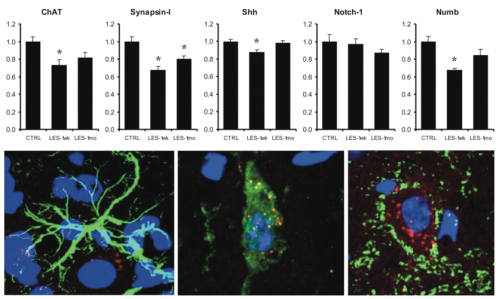
We observed that the motoneuron depletion was paralleled by a worsening of functional performances and by the down-regulation of expression of ChAT (27%), Synapsin-I (33%), Numb (32%) and Shh (12%) at one week after the lesion. Notch-1 appeared not significantly affected (Fig. 2). A significant recovery of both functional performance and protein expression levels was seen at one month after the lesion (Fig. 2). We next observed that the performance of mice at grid walk and rotarod tests strongly correlates with the expression levels of ChAT and Numb proteins in the lesioned but not in the control animals (Fig. 3). Moreover, the expression levels of ChAT and Synapsin-I, which can be considered as markers of synaptic function and plasticity, were found associated to the levels of Shh and Notch-1 (Fig. 4). Notably, the expression of the mentioned proteins has been observed in the neuronal cells, whereas glial cells appeared negative (Fig. 2). Together, these results suggest that the spontaneous recovery of locomotion could be influenced by events of synaptic plasticity and that Shh and Notch-1 could be involved in these mechanisms, whereas Numb probably modulates functional recovery by a different mechanism. Given the mentioned roles of Shh, Notch-1 and Numb, we believe that an in vivo manipulation of their signalling after lesion could represent a suitable way to improve functional recovery by modulating synaptic plasticity and/or neurogenesis. We believe that our model of SC motoneuron degeneration, induced by CTB-SAP, represents a useful tool for future studies attempting to investigate neurogenesis and/or other compensatory changes within the SC, in the presence of only neurodegenerative processes, without other microenvironmental cues such as inflammation, tissue damage, disruption of SC white matter and blood circulation.


Selected References: (back to top)
- Edgerton VR, Tillakaratne NJ, Bigbee AJ, de Leon RD, Roy RR (2004) Plasticity of the spinal neural circuitry after injury. Annu Rev Neurosci, 27:145-167.
- Gulino R, Dimartino M, Casabona A, Lombardo SA, Perciavalle V (2007) Synaptic plasticity modulates the spontaneous recovery of locomotion after spinal cord hemisection. Neurosci Res 57:148-156.
- Chen J, Leong SY, Schachner M (2005) Differential expression of cell fate determinants in neurons and glial cells of adult mouse spinal cord after compression injury. Eur J Neurosci 22:1895-1906.
- Costa RM, Honjo T, Silva AJ (2003) Learning and memory deficits in Notch mutant mice. Curr Biol 13:1348-1354.
- Presente A, Boyles RS, Serway CN, de Belle JS, Andres, AJ (2004) Notch is required for long-term memory in Drosophila. Proc Natl Acad Sci USA 101:1764-1768.