Matsuura T, Kawasaki M, Hashimoto H, Yoshimura M, Motojima Y, Saito R, Ueno H, Maruyama T, Ishikura T, Sabanai K, Mori T, Ohnishi H, Onaka T, Sakai A, Ueta Y (2016) Possible involvement of the rat hypothalamo-neurohypophysial/-spinal oxytocinergic pathways in acute nociceptive responses. J Neuroendocrinol 28(6) doi: 10.1111/jne.12396
Summary: It has been suggested that the amplification of GABAergic neurons in the inhibitory system induces the selective inhibition by Oxytocin (OXT) of excitability in the spinal cord, and the pain transmitted from the periphery to the dorsal horn of the spinal cord by this action may be attenuated at the spinal cord level. Rats were injected IT with Oxytocin-SAP (Cat. #IT-46) dissolved in saline (0.06 μg/μl), Blank-SAP (Cat. #IT-21) dissolved in saline (0.06 μg/μl), or saline. Formalin-induced acute nociception activated OXT-containing cells in both the magnocellular and parvocellular divisions of hypothalamus, and that the parvocellular division remains activated longer than the magnocellular division. Acute nociception-induced activation of the hypothalamo-neurohypophysial system caused elevation of plasma OXT levels. In addition, the OXTergic spinal pathway may be involved in pain modulation via OXTRs in the spinal cord.
Over the years, ATS has frequently been asked about Saporin’s safety for use in the lab as well as when used clinically. Residual awareness of alternate Ribosome-Inactivating Proteins (RIPs) and ‘toxins’ such as Ricin have caused some researchers new to the use of RIPs to question the belief that Saporin is safe. Unlike Type 2 RIPs (such as Ricin), Type I RIPs, like Saporin have no binding chain and consequently no means of entering the physiological space necessary for the protein to act as a toxin. The following is a review of safety in handling and potential toxicity within the human body for systemic events not related to normal research applications of Saporin conjugates, including Substance P-Saporin (SP-SAP), which is a therapeutic under development for the treatment of chronic pain.
The acute LD50 for saporin in mice (25 g) is 6.8 mg/kg;[1] that would translate in humans (75 kg) to 510 mg! A concentration of about 100 nM is the threshold to see even a vague hint of saporin toxicity. In human blood, that would correspond to 24 mg injected systemically into a person. The fermentation process to produce recombinant saporin has a titer of 2 mg/L meaning that the production broth itself contains no more than 67 nM concentration of saporin. Furthermore, the final protein concentrations from production batches of recombinant Saporin used in our drug are 4 mg/ml, meaning 6 mL of final material would need to accidentally end up in a human before the ‘hint of toxicity’ threshold would potentially be met.
The toxicology studies of SP-SAP contained within ATS’s IND prior to the current human Phase I clinical trial evaluated effects related to the intended method of administration, intrathecal local injection. SP-SAP is not expected to ever be a self-administered therapy, so the effects of gross off-target events, such as accidental auto-injection, swallowing, spillage, or immersion were not considered.
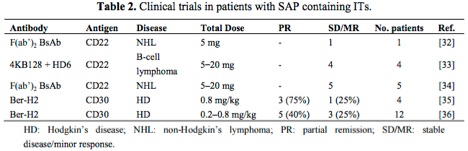
The table below[2] highlights antibody-saporin conjugates approved by the FDA for Phase I/II clinical trials in humans. The therapeutics listed below were administered intravenously and imply what the FDA accepted as non-toxic levels of saporin-based conjugates in these studies.
Looking more closely at the study by French et al.,[3] several milligrams of antibody conjugate were repeatedly injected into human patients under a FDA regulated clinical trial and peak serum levels tested, demonstrating rapid clearing of saporin from the system.
As a company that specializes in Saporin, our two-plus decades of experience working with the protein in research, preclinical, and clinical environments has taught us that with minimal standard laboratory precautions users are not at any real risk of toxic effects. Even our CSO, after 30+ years of working with Saporin exhibits undetectable levels of Saporin antibodies in his blood!
Vierck C, Yezierski R, Wiley R (2016) Pain sensitivity following loss of cholinergic basal forebrain (CBF) neurons in the rat. Neuroscience 319:23-34. doi: 10.1016/j.neuroscience.2016.01.038
Objective: There is a large amount of research on the involvement of cholinergic mechanisms on spinal transmission of pain signals, indicating that cholinergic agonists can attenuate this kind of pain. In contrast, some studies have shown affective reactions to pain are suppressed by cholinergic antagonists. The authors investigated the disagreement between reflexive and affective reactions.
Summary: Lesioned rats displayed decreased escape from thermal stimulation, as well as loss of the normal hyperalgesic effect of sound stress. Results indicate that the basal forebrain cholinergic system plays a role in central processing of pain.
Usage: Administration of 192-IgG-SAP with a 4-μg injection into the left lateral ventricle of rats. Animals were tested in temperature escape and sound stress models.
Yao Y, Echeverry S, Shi X, Yang M, Yang Q, Wang G, Chambon J, Wu Y, Fu K, De Koninck Y, Zhang J (2016) Dynamics of spinal microglia repopulation following an acute depletion. Sci Rep 6:22839. doi: 10.1038/srep22839
Summary: This study confirms that similar to microglia in the brain, spinal microglia can repopulate rapidly following elimination, which is driven essentially by a self-renewal process. To deplete microglia in spinal cords, Mac-1-SAP (Cat. #IT-06) was injected i.t. (7 μl, 1.6 μg/μl) at the level of L4-L5 in mouse. The results support the concept that microglia repopulation, whether in the brain or in the spinal cord, is the consequence of onsite resident microglia proliferation. Newly generated microglia are fully functional and are able to respond to peripheral nerve injury and contribute to the development of neuropathic pain.
Fei Y, Wang X, Chen S, Zhou Q, Zhang C, Li Y, Sun L, Zhang L (2016) Role of the RVM in descending pain regulation originating from the cerebrospinal fluid-contacting nucleus. Neurochem Res 41:1651-1661. doi: 10.1007/s11064-016-1880-6
Summary: The researchers investigated whether the CSF-contacting nucleus contributed to descending pain modulation in normal and neuropathic rats, and detected the 5-HT expression changes in both RVM and spinal dorsal cord. They also detected the possible anatomical and function correlation between the CSF-contacting nucleus and the RVM. Targeted ablation of the CSF-contacting nucleus was performed using CTB-SAP (Cat. #IT-14; 500 ng/3 μl), which was administered i.c.v. to the normal rats and rats 7 days before the CCI procedure. Based on the findings of the present study, they believe that the CSF-contacting nucleus may act as a component of descending pain regulation system. RVM, which acts as an important brain nucleus, is involved in the relay of nociceptive information between the CSF-contacting nucleus and spinal cord. Moreover, RVM 5-HT system plays a critical role in descending pain inhibition originating from the CSF-contacting nucleus.
Gritsch S, Bali K, Kuner R, Vardeh D (2016) Functional characterization of a mouse model for central post-stroke pain. Mol Pain 12:1744806916629049. doi: 10.1177/1744806916629049
Summary: While clinical evidence has pointed toward central pain pathway dysfunction in central post-stroke pain (CPSP), the underlying mechanisms have not been defined. In this work the authors created a mouse model of CPSP through lesions of the thalamic ventral posterolateral nucleus. In order to examine the role of neurokinin-1 receptor-expressing (NK1R) neurons in lamina I/III of the spinal cord in the development and maintenance of CPSP the authors administered 1 μmol intrathecal injections of SSP-SAP (Cat. #IT-11). Saporin (Cat. #PR-01) was used as a control. While the NK1R+ neurons in the spinal cord were not involved in establishing CPSP, the data indicate that sensory changes in the mice are comparable to those observed in human patients with CPSP.
La J, Feng B, Kaji K, Schwartz E, Gebhart G (2016) Roles of isolectin B4-binding afferents in colorectal mechanical nociception. Pain 157:348-354. doi: 10.1097/j.pain.0000000000000380
Summary: Primary afferent neurons are often classified as peptidergic or non-peptidergic. One characteristic of the non-peptidergic neurons is that they bind isolectin-B4. In the spinal cord these neurons terminate mainly in inner lamina II. Non-peptidergic neurons in the spinal cord have been found to be involved in various aspects of pain response. In this work the authors examined the role of non-peptidergic neurons in the viscerosensory system. Rats received 1.5 μg of intrathecal recombinant IB4-SAP (Cat. #IT-10) between the L5 and L6 vertebrae. Saporin (Cat. #PR-01) was used as a control. While IHC demonstrated that a majority of viscerosensory L6 colon DRG neurons are IB4+, they do not play a significant role in colorectal mechano-nociception.
Webster C, Caram-Salas N, Haqqani A, Thom G, Brown L, Rennie K, Yogi A, Costain W, Brunette E, Stanimirovic D (2016) Brain penetration, target engagement, and disposition of the blood-brain barrier-crossing bispecific antibody antagonist of metabotropic glutamate receptor type 1. FASEB J 30:1927-1940. doi: 10.1096/fj.201500078 PMID: 26839377
Summary: To generate a BBB-transmigrating antibody that could be reformatted to full IgG, scientists started with the BBB-crossing llama single domain antibody FC5. Standard phage display protocols were used to isolate single-chain variable fragments (scFv) from the FC5-scFv library. 6His Mouse Monoclonal antibody (Cat. #AB-213) was used to assess cell binding of scFvs of FC5 using fluorescence microvolume assay technology. An scFv that competed with FC5 binding was selected for further testing. An antibody antagonist of the metabotropic glutamate receptor-1 was fused with this scFv antibody fragment (BBB-mGluR1) and tested in an in vitro BBB model. The resulting bispecific antibody retained selective mGluR1 binding and saw a 20-fold enhanced rate of transcytosis across the BBB compared to fusion with control antibody fragment. Intravenous injection of BBB-mGluR1 had analgesic properties in a rat model of persistent inflammatory pain.
Lee J, Jeong D, Lee J, Chang W, Chang J (2016) The effect of nucleus basalis magnocellularis deep brain stimulation on memory function in a rat model of dementia. BMC Neurol 16:6. doi: 10.1186/s12883-016-0529-z
Objective: Deep brain stimulation (DBS) is the application of electrical impulses to specific parts of the brain for treating disorders such as Parkinson’s disease, chronic pain, and obsessive-compulsive disorder. This study investigated whether stimulation of brain structures associated with memory can enhance cognitive function.
Summary: Results indicate that DBS has beneficial effects on consolidation and retrieval of visuospatial memory.
Usage: The authors lesioned the basal forebrain of rats through bilateral injections totaling 5 μg of 192-IgG-SAP into the lateral ventricle. Animals then received DBS to the nucleus basalis magnocellularis and were tested in a Morris water maze task.
Harasawa I, Johansen J, Fields H, Porreca F, Meng I (2016) Alterations in the rostral ventromedial medulla after the selective ablation of μ-opioid receptor expressing neurons. Pain 157:166-173. doi: 10.1097/j.pain.0000000000000344
Summary: The rostral ventromedial medulla (RVM) has both excitatory and inhibitory control over nociceptive neurons in the medullary dorsal horn and spinal cord. Previous work has demonstrated that elimination of mu-opioid receptor-expressing neurons in the RVM reduces stress and injury-induced behavioral hypersensitivity, but the effect of losing these cells on the descending inhibitory system has not been examined. The authors administered 1.2 pmol of Dermorphin-SAP (Cat. #IT-12) to each side of the RVM of rats. Saporin (Cat. #PR-01) was used as a control. Characterization of RVM neurons in lesioned animals showed a reduction in on- and off-cells, but no change in the number of neutral cells. These data indicate that mu-opioid receptor-expressing cells in the RVM are not needed for analgesia produced by activation of RVM neurons.