Contributed by Dr. Patrick O. Kanold, Dept Neurobiology, Harvard Medical School, Boston, MA 02115.
Dr. Kanold summarizes his work with ME20.4-SAP. A complete report was published in Science 301:521-525.
The processing of information in the visual system happens in multiple stages. Retinal neurons in the eye connect to neurons in the lateral geniculate nucleus (LGN), that in turn connect to neurons in the visual cortex. The axonal fibers grow to connect these regions during fetal development. For example, fibers from the LGN grow into visual cortex. Once these LGN axons reach their target area in visual cortex, they start to form synapses with neurons in the area. The axons also start to segregate into the ocular dominance columns (ODCs), stripes of cortex that are connected exclusively to the left eye or to the right eye. The prevailing hypothesis about the formation of ODCs is that they are shaped by activity-dependent competition between afferents from each eye and by selective growth and pruning of axonal projections as a means of refinement. The factors and circuits mediating the competition remain to be elucidated.
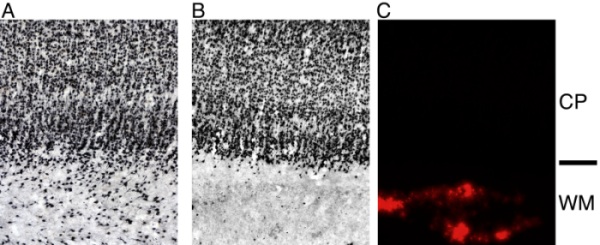
During this period of connecting and remodeling, a peculiar group of neurons is present in the developing brain: the subplate neurons. In contrast to other neuronal structures in the brain, this is a transient population of neurons. Subplate neurons are the first to mature in the developing cortex but then later die during further development. The axons of the subplate neurons reach into the developing cortex at a very early stage. Later, on the way to the cortex, thalamic axons pass through the subplate and wait there before they grow into the developing cortex. In part because of the brief life span of subplate neurons, very little is known about their function.
However, subplate neurons are in a key, intermediate position to control the flow of information into the developing cortex when first spontaneous (prenatal) and then visual (postnatal) activity are present in visual cortex. The fact that ODCs emerge even before the onset of patterned visual experience (1, 2), poses the question how subplate neurons influence the functional development of cortex. The subplate forms a transient circuit that is required for the development of axonal projections between thalamus and cerebral cortex. When subplate neurons are ablated after the arrival of thalamic inputs to layer 4 (P7-P10), ODCs do not form despite the robust presence of thalamic axons (3). We found that in addition to defects in the anatomical organization of visual cortex, late subplate ablation [using kainic acid or ME20.4-SAP (Cat. #IT-15)] leads to profound functional deficits (4). Following the ablation, functional orientation maps are disorganized and visual responses are weak (4). Extracellular recordings in vivo showed that the orientation tuning of single neurons in visual cortex was weak and that these neurons were only weakly responsive to visual stimuli. In vitro slice recordings and in vivo current-source density measurements show that subplate ablation results in reduced efficacy of thalamocortical synaptic transmission consistent with the lower expression of GluR1 mRNA expression in layer 4 of the ablated area (4).
We conclude that subplate neurons and their connections are needed not only for the anatomical segregation of thalamic inputs into ODCs, but also for key steps in functional synaptic maturation and remodeling involved in creating the highly tuned responses of adult cortical neurons in all layers.
These observations are noteworthy because the thalamic axons from the LGN are all still present within cortical layer 4, however, they do not strengthen or refine themselves into adult patterns of connections (3, 4). In contrast, transmission from layer 4 to layer 2/3 is unaffected by subplate ablation (4) indicating that, apparently, cortex can continue to mature and develop even when not functionally activated by thalamic inputs.
References
- Crowley JC, Katz LC (2000) Early development of ocular dominance columns. Science 290(5495):1321-1324.
- Crair MC, Horton JC, Antonini A, Stryker MP (2001) Emergence of ocular dominance columns in cat visual cortex by 2 weeks of age. J Comp Neurol 430(2):235-249.
- Ghosh A, Shatz CJ (1992) Involvement of subplate neurons in the formation of ocular dominance columns. Science 255(5050):1441-1443.
- Kanold PO, Kara P, Reid RC, Shatz CJ (2003) Role of subplate neurons in functional maturation of visual cortical columns. Science 301(5632):521-525, 2003.